




 Production of Thyroid Hormone Part One by Jeffrey Dach MD
Production of Thyroid Hormone Part One by Jeffrey Dach MD
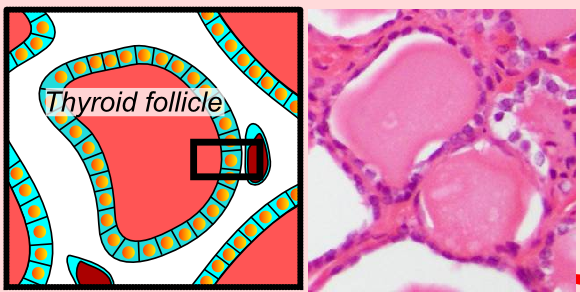
Above image: At left is schematic of the thyroid follicle with inset box showing magnified view of thyroid hormone synthesis (next image). Image at right is typical microscopic view of thyroid follicle showing central round areas of pink colloid surrounded by lining of follicular cells (normal gland).
Magnified Cross Section of Thyrocyte and Follicular Lumen:

Above Image : Magnified view of inset box: Diagram of Thyroid Hormone Synthesis showing active uptake of Iodine from the blood stream by the Sodium-Iodine-Symporter (Na/I Symporter) at the basal membrane of the thyrocyte (purple arrow at far right). The iodine travels through the thyrocyte to the left and is pumped into the follicular lumen by the Pendrin complex at the apical membrane.(purple arrow left). Upper area of diagram, separately, thyroglobulin protein is manufactured in the endoplasmic reticulum and secreted into the follicular lumen by exocytosis (left pink arrow). This thyroglobulin in the follicular lumen is called colloid. The thyroglobulin is then Iodinated by the thyroperoxidase enzyme (see next image panel zoomed image)
Iodination of Thyroglobulin by Thyroperoxidase, Apical Membrane and Follicular Lumen:

Above Image: showing oxidation of Iodine by the thyroperoxidase enzyme on surface of villi on inner apical membrane of thyrocytes. (purple box). The activated iodine will then spontaneously combine with Tryrosine residues on the thyroglobulin. The Tryrosine molecules are conjugated together in the next step to form thyroxine (thyroid hormone), which is then secreted back into the blood stream by the thyrocytes.
Hashimoto’s Thyroiditis:
Antibodies to the Thyroperoxidase enzyme prevent it from functioning properly, resulting in decreased or absent ability to iodinate the thyroglobulin. This is called “inability to organify iodine”, and explains the low iodine content of the thyroid gland in Hashimotos patients, and the rapid washout of iodine with the perchlorate discharge test. This also explains how the thyroid gland in Hashimoto’s is more sensitive to “Iodine Blockade” described by Wolf, the suppressive effect of Iodine on thyroid function.
A second antibody against thyroglobulin may also be present. Additional antibodies against the TSH receptor, and against Pendrin have also been discovered. More antibodies may be discovered in the future.
Methimazole-antithyroid medication
Methimazole works by blocking the function of the thyroidperoxidase enzyme, preventing iodination of thyroglobulin. (9) Selenium analoges of Methimazole, still under development, may be preferable because they protect the thyroid from oxidation.(10)
Genetic Mutation in Thyroperoxidase
In people who have a genetic mutation in the thyroperoxidase enzyme, their thyroid glands cannot organify iodine. The iodide in their thyroid gland cannot be oxidized and/or bound to the thyroglobulin protein.(8)
The role of selenium and hydrogen peroxide
The thyroid gland has a high selenium content due to selenoproteins (glutathione peroxidases) which protect the cells from oxidative damage associated with hydrogen peroxide production needed for thyroid hormone biosynthesis. (11)
Iodination of thyroglobulin is the key step of thyroid hormone biosynthesis. It is catalyzed by thyroid peroxidase and occurs within the follicular space at the apical plasma membrane.(12-14) Hydrogen peroxide generating enzymes are also found at this same location, at the villous apical membrane of the thyrocyte just within the follicular lumen.(12-14) Both enzyme systems are needed for the organification of iodine to throglobulin proteins. Various thyroid pathologies, including hashimotos thyroiditis can be explained by overproduction and lack of degradation of hydrogen peroxide (H2O2) causing damage to the thyrocyte structures. (15,16)
Iodine Intake Inhibits H202 Generation-Wolf Chaikoff Effect
The “Wolff-Chaikoff” effect is the inhibition of H2O2 generation caused by iodine itself, The intake of dietary Iodine, itself, will limit the oxidation and binding of iodide to thyroglobulin because of the reduced availability of hydrogen peroxide at the apical membrane. (17) The Wolff-Chaikoff effect is the inhibition by iodide of its own organification. (17) Since the organification of iodine is already reduced in Hashimotos thyroiditis, these patients are more sensitive to the inhibitory effect of iodine intake on thyroid hormone production. This will be discussed later in various studies which misinterpret the inhibitory effect of Iodine as a worsening in Hashimotos thyroiditis.
Watch this animated video of thyroid hormone production:
link to this article: http://wp.me/p3gFbV-1gz
Jeffrey Dach MD
7450 Griffin Road, Suite 190
Davie, Fl 33314
954-792-4663
Articles with related interest:
Hashimotos Thyroiditis, Manic Depression, Psychosis and Psychiatric Manifestations
Iodine and Hashimotos Thyroid Disease
Hashimotos Thyroid Disease and Molecular Mimicry
Hashimotos Thyroiditis and Selenium Part One
Hashimotos, Selenium and Iodine, Part Two
Hashimotos and Iodine Part Three
Selenium and the Thyroid More Good News
Links and References
SYNTHESIS OF THYROID HORMONES, thyroperoxidase iodine
1) www.youtube.com/embed/
Thyroid Hormone Production video on You Tube
2) Image for Synthesis of Thyroid Hormone: Image file on WIkimedia commons: http://commons.wikimedia.org/
3) https://www.inkling.com/read/
Synthesis of Thyroid Hormones Thyroid hormones are made by iodinating tyrosine residues on thyroglobulin and are stored as part of thyroglobulin molecules in thyroid follicles.
Synthesis of thyroid hormones. Chapter 48, “SYNTHESIS OF THYROID HORMONES” in: Walter F., PhD. Boron (2003) Medical Physiology: A Cellular And Molecular Approach, Elsevier/Saunders, pp. 1,300 ISBN: 1-4160-2328-3.
4) http://www.medical-
Central:Thyroid hormones (T4 and T3) are produced by the follicular cells of the thyroid gland and are regulated by TSH made by the thyrotropes of the anterior pituitary gland. The effects of T4 in vivo are mediated via T3 (T4 is converted to T3 in target tissues). T3 is 3- to 5- fold more active than T4.
Thyroxine (3,5,3′,5′-tetraiodothyronine) is produced by follicular cells of the thyroid gland. It is produced as the precursor thyroglobulin (this is not the same as TBG), which is cleaved by enzymes to produce active T4.
The steps in this process are as follows:
The Na+/I- symporter transports two sodium ions across the basement membrane of the follicular cells along with an iodine ion. This is a secondary active transporter that utilises the concentration gradient of Na+ to move I- against its concentration gradient.
I- is moved across the apical membrane into the colloid of the follicle.
Thyroperoxidase oxidises two I- to form I2. Iodide is non-reactive, and only the more reactive iodine is required for the next step.
The thyroperoxidase iodinates the tyrosyl residues of the thyroglobulin within the colloid. The thyroglobulin was synthesised in the ER of the follicular cell and secreted into the colloid.
Thyroid-stimulating hormone (TSH) released from the pituitary gland binds the TSH receptor ( a Gs protein-coupled receptor) on the basolateral membrane of the cell and stimulates the endocytosis of the colloid.
The endocytosed vesicles fuse with the lysosomes of the follicular cell. The lysosomal enzymes cleave the T4 from the iodinated thyroglobulin.
These vesicles are then exocytosed, releasing the thyroid hormones.
Thyroxine is produced by attaching iodine atoms to the ring structures of tyrosine molecules. Thyroxine (T4) contains four iodine atoms. Triiodothyronine (T3) is identical to T4, but it has one less iodine atom per molecule.
Iodide is actively absorbed from the bloodstream by a process called iodide trapping. In this process, sodium is cotransported with iodide from the basolateral side of the membrane into the cell and then concentrated in the thyroid follicles to about thirty times its concentration in the blood. Via a reaction with the enzyme thyroperoxidase, iodine is bound to tyrosine residues in the thyroglobulin molecules, forming monoiodotyrosine (MIT) and diiodotyrosine (DIT). Linking two moieties of DIT produces thyroxine. Combining one particle of MIT and one particle of DIT produces triiodothyronine.
DIT + MIT → r-T3 (biologically inactive)
MIT + DIT → triiodothyronine (usually referred to as T3)
DIT + DIT → thyroxine (referred to as T4)
Proteases digest iodinated thyroglobulin, releasing the hormones T4 and T3, the biologically active agents central to metabolic regulation.
Peripheral:
Thyroxine is believed to be a prohormone and a reservoir for the most active and main thyroid hormone T3. T4 is converted as required in the tissues by iodothyronine deiodinase. Deficiency of deiodinase can mimic an iodine deficiency. T3 is more active than T4 and is the final form of the hormone, though it is present in less quantity than T4. See more at: http://www.medical-
——————————
5) http://www.ncbi.nlm.nih.gov/
J Biol Chem. 1984 Jan 10;259(1):359-64.
Iodination and oxidation of thyroglobulin catalyzed by thyroid peroxidase. Nakamura M, Yamazaki I, Nakagawa H, Ohtaki S, Ui N.
Thyroid peroxidase plays catalytic roles in two steps of the biosynthesis of thyroxine:
(a) the iodination of tyrosine residues of thyroglobulin and
(b) the coupling of diiodotyrosine to produce thyroxine
thyroid peroxidase oxidizes iodide to atomic iodine (I) or iodinium (I+).
The “organification of iodine,” the incorporation of iodine into thyroglobulin for the production of thyroid hormone,
is nonspecific; that is, there is no TPO-bound intermediate, but iodination occurs via reactive iodine species released from TPO.[4]
The chemical reactions catalyzed by thyroid peroxidase occur on the outer apical membrane surface and are mediated by hydrogen peroxide.
6) http://www.ncbi.nlm.nih.gov/
Front Biosci (Landmark Ed). 2011 Jan 1;16:783-802.
Thyroperoxidase, thyroglobulin, Na(+)/I(-) symporter, pendrin in thyroid autoimmunity.Czarnocka B.
——————————
Location of thyro peroxidase
7) http://www.ncbi.nlm.nih.gov/
CYTOCHEMICAL LOCALIZATION OF ENDOGENOUS PEROXIDASE IN THYROID FOLLICULAR CELLS
Judy M. Strum and Morris J. Karnovsky
Peroxidase activity is found in thyroid follicular cells in the following sites:apical vedicles, associated with the external surfaces of the microvilli that project apically from the cell into the colloid.
In keeping with the radioautographic evidence of others and the postulated role of thyroid peroxidase in iodination, it is suggested that the microvillous apical cell border is the major site where iodination occurs.
Mutation in the Throperoxidase Gene- cannot organify iodine
8) http://www.ncbi.nlm.nih.gov/
J Endocrinol. 2002 Mar;172(3):627-35.
Mutation analysis of thyroid peroxidase gene in Chinese patients with total iodide organification defect: identification of five novel mutations. Wu JY, Shu SG, Yang CF, Lee CC, Tsai FJ.
Total iodide organification defect (TIOD), where the iodide in the thyroid gland cannot be oxidized and/or bound to the protein, is caused by a defect in the thyroid peroxidase (TPO) gene. Single strand conformation polymorphism analysis was used to screen for mutations in the TPO gene from five unrelated TIOD patients in Taiwan, and five novel mutations were detected. Three of these were frameshift mutations: a single T insertion between nucleotide position 2268 and 2269 (c.2268-2269 insT) in exon 13 and two single C deletions at nucleotide positions 843 (c.843 delC) and 2413 (c.2413 delC) in exon 8 and 14 respectively. The other two were single nucleotide substitutions (c.G1477>A and c.G2386>T) located in exons 9 and 13 respectively. While the former would result in amino acid substitution (Gly493Ser) in the highly conserved region of the TPO polypeptide, the latter would result in either amino acid substitution (Asp796Tyr) or alternative splicing. Of those identified TPO mutations, c.2268-2269 insT was most prevalent and was detected as heterozygous in all but one TIOD patients. All five TIOD patients investigated in this study were compound heterozygous. The method presented in this study could be used for carrier assessment and mutation analysis of newly identified TIOD patients.
9) http://www.ncbi.nlm.nih.gov/pubmed/947933
J Clin Endocrinol Metab. 1976 Jul;43(1):152-8.
Effect of antithyroid agents 6-propyl-2-thiouracil and 1-mehtyl-2-mercaptoimidazole on human thyroid iodine peroxidase.
Nagasaka A, Hidaka H.
The mechanism of inhibition of human thyroid iodide peroxidase (TPO) by 6-propyl-2-thiouracil (PTU) and 1-methyl-2-mercaptoimidazole (MMI) used in the therapy of hyperthyroid patients was studied in vitro. The inhibition of TPO by MMI was not restored either by dialysis or by dilution, but the inhibition by PTU was restored by both treatments. PTU interacted directly with the product of TPO action (oxidized iodide) in the reaction mixture without significantly affecting TPO activity. MMI interacted directly with TPO and inhibited enzyme activity, rather than interacting with the product (oxidized iodide). The inhibition was irreversible with MMI, but reversible with PTU. The concentrations of PTU and MMI producing 50% inhibition of TPO were 2 x 10-6m and 8 x 10-7m, respectively, 2-Mercaptoimidazole inhibited TPO reversibly but 1-methylimidazole and imidazole did not. Both the methyl and mercaptoresidues in MMI moiety are thought to be essential to its irreversible inhibition of TPO. The in vivo effect of MMI and PTU on TPO activity was also studied. TPO activities in the thyroid homogenate of rats to which MMI (2 mg per rat) or PTU (10 mg per rat) had been administered intraperitoneally were determined before and after dialysis against buffer. TPO activity in the PTU treated thyroid homogenate was significantly lower than that in the control before dialysis, but the activity was restored to the control value after dialysis. On the contrary, TPO activity in the MMI treated thyroid homogenate was significantly lower than that in the control and was not affected by dialysis. These data may explain why MMI is a more potent inhibitor of iodination than PTU and may fit the clinical results observed when hyperthyroid patients are treated with these agents.
10) http://www.ncbi.nlm.nih.gov/pubmed/18357551
Chem Biodivers. 2008 Mar;5(3):414-39. doi:
Selenium analogues of antithyroid drugs–recent developments.
Roy G, Mugesh G. Thyroxine (T4), the main secretory hormone of the thyroid gland, is produced on thyroglobulin by thyroid peroxidase (TPO)/H(2)O(2)/iodide system and deiodinated to its active form (T3) by a selenocysteine-containing enzyme, iodothyronine deiodinase (ID). The activation of thyroid-stimulating hormone (TSH) receptor by auto-antibodies leads to ‘hyperthyroidism’, a life-threatening disease which is treated by antithyroid drugs such as 6-propyl-2-thiouracil (PTU) and methimazole (MMI). The present review describes the biological activities of a number of S/Se derivatives bearing the methimazole pharmacophore. It is shown that the isosteric substitutions in the existing drugs lead to compounds that can effectively and reversibly inhibit the heme-containing lactoperoxidase (LPO). In contrast to methimazole, the selenium analogue, MSeI, does not interfere with the enzyme directly, but it inhibits LPO by reducing the H(2)O(2) that is required for the oxidation of the Fe-center in LPO.
———————————————————————-
Selenoproteins in the thyroid
11) http://www.ncbi.nlm.nih.gov/
Biol Chem. 2007 Oct;388(10):1053-9.
Selenoproteins of the thyroid gland: expression, localization and possible function of glutathione peroxidase Schmutzler C, Mentrup B, Schomburg L, Hoang-Vu C, Herzog V, Köhrle J.
The thyroid gland has an exceptionally high selenium content, even during selenium deficiency. At least 11 selenoproteins are expressed, which may be involved in the protection of the gland against the high amounts of H2O2 produced during thyroid hormone biosynthesis. As determined here by in situ hybridization and Northern blotting experiments, glutathione peroxidases (GPx) 1 and 4 and selenoprotein P were moderately expressed, occurring selectively in the follicular cells and in leukocytes of germinal follicles of thyroids affected by Hashimoto’s thyroiditis. Selenoprotein 15 was only marginally expressed and distributed over all cell types. GPx3 mRNA was exclusively localized to the thyrocytes, showed the highest expression levels and was down-regulated in 5 of 6 thyroid cancer samples as compared to matched normal controls. GPx3 could be extracted from thyroidal colloid by incubation with 0.5% sodium dodecyl sulfate indicating that this enzyme is (i) secreted into the follicular lumen and (ii) loosely attached to the colloidal thyroglobulin. These findings are consistent with a role of selenoproteins in the protection of the thyroid from possible damage by H2O2. Particularly, GPx3 might use excess H2O2 and catalyze the polymerization of thyroglobulin to the highly cross-linked storage form present in the colloid.
12) http://www.ncbi.nlm.nih.gov/
Antioxid Redox Signal. 2008 Sep;10(9):1577-92. doi: 10.1089/ars.2008.2054.
Peroxides and peroxide-degrading enzymes in the thyroid.
Schweizer U, Chiu J, Köhrle J.
Iodination of thyroglobulin is the key step of thyroid hormone biosynthesis. It is catalyzed by thyroid peroxidase and occurs within the follicular space at the apical plasma membrane. Hydrogen peroxide produced by thyrocytes as an oxidant for iodide may compromise cellular and genomic integrity of the surrounding cells, unless these are sufficiently protected by peroxidases. Thus, peroxidases play two opposing roles in thyroid biology. Both aspects of peroxide biology in the thyroid are separated in space and time and respond to the different physiological states of the thyrocytes. Redox-protective peroxidases in the thyroid are peroxiredoxins, glutathione peroxidases, and catalase. Glutathione peroxidases are selenoenzymes, whereas selenium-independent peroxiredoxins are functionally linked to the selenoenzymes of the thioredoxin reductase family through their thioredoxin cofactors. Thus, selenium impacts directly and indirectly on protective enzymes in the thyroid, a link that has been supported by animal experiments and clinical observations. In view of this relationship, it is remarkable that rather little is known about selenoprotein expression and their potential functional roles in the thyroid. Moreover, selenium-dependent and -independent peroxidases have rarely been examined in the same studies. Therefore, we review the relevant literature and present expression data of both selenium-dependent and -independent peroxidases in the murine thyroid.
13) http://www.ncbi.nlm.nih.gov/
Histochemistry. 1985;82(3):263-8.
Cytochemical localization of peroxidase and hydrogen-peroxide-producing NAD(P)H-oxidase in thyroid follicular cells of propylthiouracil-treated rats.
Mizukami Y, Matsubara F, Matsukawa S.
Abstract The distribution of endogenous peroxidase and hydrogen-peroxide-producing NAD(P)H-oxidase, which are essential enzymes for the iodination of thyroglobulin, was cytochemically determined in the thyroid follicular cells of propylthiouracil (PTU)-treated rats. Peroxidase activity was determined using the diaminobenzidine technique. The presence of NAD(P)H-oxidase was determined using H2O2 generated by the enzyme; the reaction requires NAD(P)H as a substrate and cerous ions for the formation of an electron-dense precipitate. Peroxidase activity was found in the developed rough endoplasmic reticulum (rER) and Golgi apparatus, but it was also associated with the apical plasma membrane; NAD(P)H-oxidase activity was localized on the apical plasma membrane. The presence of both enzymes on the apical plasma membrane implies that the iodination of thyroglobulin occurs at the apical surface of the follicular cell in the TSH-stimulated state which follows PTU treatment.
14) http://www.ncbi.nlm.nih.gov/
Tissue Cell. 1985;17(6):889-900.
Cytochemical localization of hydrogen peroxide generating sites in the rat thyroid gland. Labato MA, Briggs RT.
Abstract Sites of H2O2 generation in lightly prefixed, intact thyroid follicles were studied by two cytochemical reactions: peroxidase-dependent DAB oxidation and cerium precipitation. In both cases reaction product accumulated on the apical surface of the follicle cell at the membrane-colloid interface. The former reaction was inhibited by the peroxidase inhibitor, aminotriazole; both reactions were blocked by the presence of catalase. NADH in the medium slightly increased the amount of cerium precipitation. The ferricyanide technique for oxidoreductase activity was also applied; reaction product again was associated with the apical surface. These results strongly imply that the follicle cells have a NADH oxidizing system generating H2O2 at the apical plasma membrane.
15) http://www.ncbi.nlm.nih.gov/
J Clin Endocrinol Metab. 2007 Oct;92(10):3764-73. Epub 2007 Jul 31.
Roles of hydrogen peroxide in thyroid physiology and disease.
Song Y, Driessens N, Costa M, De Deken X, Detours V, Corvilain B, Maenhaut C, Miot F, Van Sande J, Many MC, Dumont JE.
The long-lived thyroid cell generates, for the synthesis of thyroid hormones, important amounts of H2O2 that are toxic in other cell types. This review analyzes the protection mechanisms of the cell and the pathological consequences of disorders of this system.
EVIDENCE ACQUISITION:The literature on H2O2 generation and disposal, thyroid hormone synthesis, and their control in the human thyroid is analyzed.
EVIDENCE SYNTHESIS:In humans, H2O2 production by dual-oxidases and consequently thyroid hormone synthesis by thyroperoxidase are controlled by the phospholipase C-Ca2+-diacylglycerol arm of TSH receptor action. H2O2 in various cell types, and presumably in thyroid cells, is a signal, a mitogen, a mutagen, a carcinogen, and a killer. The various protection mechanisms of the thyroid cell against H2O2 are analyzed. They include the separation of the generating enzymes (dual-oxidases), their coupling to thyroperoxidase in a proposed complex, the thyroxisome, and H2O2 degradation systems.
CONCLUSIONS:It is proposed that various pathologies can be explained, at least in part, by overproduction and lack of degradation of H2O2 (tumorigenesis, myxedematous cretinism, and thyroiditis) and by failure of the H2O2 generation or its positive control system (congenital hypothyroidism).
16) http://www.ncbi.nlm.nih.gov/
Exp Biol Med (Maywood). 2010 Apr;235(4):424-33. Dual oxidase, hydrogen peroxide and thyroid diseases. Ohye H, Sugawara M.
The thyroid gland is a unique endocrine organ that requires hydrogen peroxide (H(2)O(2)) for thyroid hormone formation. The molecule for H(2)O(2) production in the thyroid gland has been known as dual oxidase 2 (DUOX2). Recently, NADPH oxidase 4 (NOX4), a homolog of the NOX family, was added as a new intracellular source of reactive oxygen species (ROS) in the human thyroid gland. This review focuses on the recent progress of the DUOX system and its possible contribution to human thyroid diseases. Also, we discuss human thyroid diseases related to abnormal H(2)O(2) generation. The DUOX molecule contains peroxidase-like and NADPH oxidase-like domains. Human thyroid gland also contains DUOX1 that shares 83% similarity with the DUOX2 gene. However, thyroid DUOX1 protein appears to play a minor role in H(2)O(2) production. DUOX proteins require DUOX maturation or activation factors (DUOXA1 or 2) for proper translocation of DUOX from the endoplasmic reticulum to the apical plasma membrane, where H(2)O(2) production takes place. Thyroid cells contain antioxidants to protect cells from the H(2)O(2)-mediated oxidative damage. Loss of this balance may result in thyroid cell dysfunction and thyroid diseases. Mutation of either DUOX2 or DUOXA2 gene is a newly recognized cause of hypothyroidism due to insufficient H(2)O(2) production. Papillary thyroid carcinoma, the most common thyroid cancer, is closely linked to the increased ROS production by NOX4. Hashimoto’s thyroiditis, a common autoimmune thyroid disease in women, becomes conspicuous when iodide intake increases. This phenomenon may be explained by the abnormality of iodide-induced H(2)O(2) or other ROS in susceptible individuals. Discovery of DUOX proteins and NOX4 provides us with valuable tools for a better understanding of pathophysiology of prevalent thyroid diseases.
17) http://www.ncbi.nlm.nih.gov/
Biochem Biophys Res Commun. 1988 Aug 15;154(3):1287-92.
Inhibition by iodide of iodide binding to proteins: the “Wolff-Chaikoff” effect is caused by inhibition of H2O2 generation. Corvilain B, Van Sande J, Dumont JE.
H2O2 generation is limiting the oxidation and binding to proteins of iodide. In dog thyroid slices thyrotropin and carbamylcholine greatly enhance protein iodination and H2O2 generation. The action of thyrotropin is mimicked by dibutyryl cyclic AMP and forskolin which suggests that it is mediated by cyclic AMP. The action of carbamylcholine was mimicked by ionomycin and by phorbol myristate ester. This suggests that the effect of carbamylcholine is mediated by the two intracellular signals generated by the Ca++ phosphatidylinositol cascade: Ca++ and diacylglycerol. The Wolff-Chaikoff effect is the inhibition by iodide of its own organification. In dog thyroid slices, iodide greatly inhibited H2O2 generation stimulated by thyrotropin and by carbamylcholine. Iodide decreased the production of intracellular signals induced by TSH and carbamylcholine but it also inhibited the action of probes of these intracellular signals (dibutyryl cAMP, forskolin, ionomycin, phorbol-myristate ester) on the H2O2 generating system itself. These effects were suppressed by methimazole an inhibitor of iodide oxidation.
link to this article: http://wp.me/p3gFbV-1gz
Jeffrey Dach MD
7450 Griffin Road, Suite 190
Davie, Fl 33314
954-792-4663
www.jeffreydach.com
www.drdach.com
www.naturalmedicine101.com
www.bioidenticalhormones101.com
www.truemedmd.com
Click Here for: Dr Dach’s Online Store for Pure Encapsulations Supplements
Click Here for: Dr Dach’s Online Store for Nature’s Sunshine Supplements
Web Site and Discussion Board Links:
jdach1.typepad.com/blog/
disc.yourwebapps.com/Indices/244124.html
disc.yourwebapps.com/Indices/244066.html
disc.yourwebapps.com/Indices/244067.html
disc.yourwebapps.com/Indices/244161.html
disc.yourwebapps.com/Indices/244163.html
Disclaimer click here: www.drdach.com/wst_page20.html
The reader is advised to discuss the comments on these pages with his/her personal physicians and to only act upon the advice of his/her personal physician. Also note that concerning an answer which appears as an electronically posted question, I am NOT creating a physician — patient relationship. Although identities will remain confidential as much as possible, as I can not control the media, I can not take responsibility for any breaches of confidentiality that may occur.
Copyright (c) 2014 Jeffrey Dach MD All Rights Reserved. This article may be reproduced on the internet without permission, provided there is a link to this page and proper credit is given.
FAIR USE NOTICE: This site contains copyrighted material the use of which has not always been specifically authorized by the copyright owner. We are making such material available in our efforts to advance understanding of issues of significance. We believe this constitutes a ‘fair use’ of any such copyrighted material as provided for in section 107 of the US Copyright Law. In accordance with Title 17 U.S.C. Section 107, the material on this site is distributed without profit to those who have expressed a prior interest in receiving the included information for research and educational purposes.



Jeffrey Dach MD New Thyroid Book Stop the Thyroid Madness Two - Jeffrey Dach MD April 7, 2015 at 3:44 PM
[…] Production of Thyroid Hormones […]